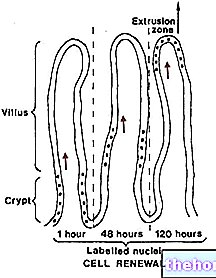
Definisi
Enzim adalah protein yang dihasilkan dalam sel tumbuhan dan haiwan, yang bertindak sebagai pemangkin mempercepat tindak balas biologi tanpa diubah suai.
Enzim berfungsi dengan menggabungkan dengan bahan tertentu untuk mengubahnya menjadi bahan yang berbeza; contoh klasik diberikan oleh enzim pencernaan yang terdapat dalam air liur, perut, pankreas dan usus kecil, yang melakukan fungsi penting dalam pencernaan dan membantu memecah makanan menjadi komponen asas, yang kemudian dapat diserap dan digunakan oleh tubuh, diproses oleh enzim lain atau dikumuhkan sebagai sampah.
Setiap enzim mempunyai peranan khusus: yang memecah lemak, misalnya, tidak bertindak terhadap protein atau karbohidrat. Enzim sangat mustahak untuk kesejahteraan organisma, kekurangan enzim tunggal boleh menyebabkan masalah serius.Contoh yang terkenal ialah fenilketonuria (PKU), penyakit yang dicirikan oleh ketidakmampuan untuk memetabolisme asid amino penting, fenilalanin, yang pengumpulannya dapat menyebabkan kecacatan fizikal dan penyakit mental.
Kajian biokimia
Enzim adalah protein tertentu yang mempunyai ciri-ciri pemangkin biologi, iaitu, mereka mempunyai keupayaan untuk menguraikan tenaga pengaktifan (Eatt) suatu tindak balas, mengubah jalannya sehingga proses kinetik yang lambat lebih cepat.
Enzim meningkatkan kinetik tindak balas termodinamik yang mungkin dan, tidak seperti pemangkin, mereka lebih kurang spesifik: oleh itu mereka mempunyai kekhususan substrat.
Enzim tidak terlibat dalam stoikiometri tindak balas: untuk ini berlaku, tapak pemangkin akhir mestilah sama dengan yang awal.
Dalam tindakan pemangkin hampir selalu ada fasa lambat yang menentukan kelajuan proses.
Apabila kita bercakap mengenai enzim, tidak betul untuk membincangkan reaksi keseimbangan, sebaliknya, kita bercakap mengenai keadaan mantap (keadaan di mana metabolit tertentu terbentuk dan dimakan secara berterusan, mengekalkan kepekatannya hampir berterusan dari masa ke masa). Produk tindak balas yang dikatalisis oleh satu enzim biasanya merupakan reaktan untuk tindak balas berikutnya, dikatalisis oleh enzim lain, dan seterusnya.
Proses yang dikatalisis oleh enzim biasanya terdiri daripada urutan tindak balas.
Reaksi generik yang dikatalisis oleh enzim (E) dapat diringkaskan sebagai berikut:

E adalah enzim
S adalah substrat;
ES mewakili penambahan antara enzim dan substrat;
P adalah produk;
K ialah pemalar kadar tindak balas.
Enzim generik (E) bergabung dengan substrat (S) untuk membentuk adduct (ES) dengan pemalar laju K1; ia dapat memisahkan kembali ke E + S, dengan pemalar laju K2, atau, (jika "hidup" cukup lama ) boleh terus membentuk P dengan pemalar kelajuan K3.
Produk (P) pada gilirannya dapat bergabung semula dengan enzim dan mereformasikan aduk dengan pemalar kadar K4.
Apabila enzim dan substrat dicampurkan, terdapat pecahan masa di mana pertemuan antara kedua spesies belum terjadi: iaitu, terdapat selang waktu yang sangat pendek (yang bergantung pada reaksi) di mana enzim dan substrat mempunyai belum dipenuhi; selepas tempoh ini, enzim dan substrat bersentuhan dalam kuantiti yang meningkat dan adduct ES terbentuk. Selepas itu, enzim bertindak pada substrat dan produk dilepaskan. Kemudian dapat dikatakan bahawa c "adalah selang waktu awal di mana kepekatan adduct ES tidak dapat ditentukan; setelah tempoh ini, diandaikan bahawa keadaan stabil ditetapkan, iaitu, kecepatan proses yang membawa kepada memperoleh adduct adalah sama dengan kecepatan proses yang menyebabkan kehancuran adduct.
Pemalar Michaelis-Menten (KM) adalah pemalar keseimbangan (disebut keseimbangan pertama yang dijelaskan di atas); dapat dikatakan, dengan pendekatan yang baik (kerana K3 juga harus dipertimbangkan), bahawa KM diwakili oleh nisbah antara pemalar kinetik K2 dan K1 (merujuk kepada pemusnahan dan pembentukan ES adduct dalam keseimbangan pertama yang dijelaskan di atas) .
Melalui pemalar Michaelis-Menten, kita mempunyai "petunjuk pertalian antara enzim dan substrat: jika KM kecil c" adalah "pertalian tinggi antara enzim dan substrat, maka penambahan ES stabil.
Enzim tertakluk kepada peraturan (atau modulasi).
Pada masa lalu terdapat perbincangan mengenai modulasi negatif, yaitu, penghambatan kemampuan katalitik enzim tetapi, ada juga modulasi positif, yaitu, ada spesies yang mampu meningkatkan kemampuan pemangkin enzim.
Terdapat 4 jenis penghambatan (diperoleh dari pendekatan yang dibuat pada model untuk memadankan data eksperimen dengan persamaan matematik):
- perencatan kompetitif
- penghambatan tidak kompetitif
- perencatan tidak kompetitif
- perencatan kompetitif
Kami bercakap mengenai perencatan kompetitif apabila molekul (perencat) mampu bersaing dengan substrat. Untuk kesamaan struktur, perencat dapat bertindak balas menggantikan substrat, oleh itu terminologi "perencatan kompetitif". Kebarangkalian enzim mengikat pada perencat atau substrat bergantung pada kepekatan kedua-duanya dan pertaliannya dengan enzim; oleh itu kadar tindak balas bergantung kepada faktor-faktor ini.
Untuk mendapatkan kadar tindak balas yang sama seperti tanpa kehadiran perencat, perlu mempunyai kepekatan substrat yang lebih tinggi.
Secara eksperimen ditunjukkan bahawa, dengan adanya perencat, pemalar Michaelis-Menten meningkat.
Sebaliknya, penghambatan "tidak kompetitif, interaksi antara molekul yang seharusnya berfungsi sebagai modulator (perencat positif atau negatif) dan enzim", berlaku di laman web yang berbeza dengan yang di mana interaksi berlaku antara enzim dan substrat; oleh itu kita bercakap mengenai modulasi alosterik (dari bahasa Yunani allosteros → laman web lain).
Sekiranya perencat mengikat enzim, ia dapat menyebabkan perubahan struktur enzim dan, akibatnya, dapat menurunkan kecekapan dengan mana substrat mengikat enzim.
Dalam proses jenis ini, pemalar Michaelis-Menten tetap berterusan kerana nilai ini bergantung pada keseimbangan antara enzim dan substrat dan, walaupun terdapat perencat, keseimbangan ini tidak berubah.
Fenomena perencatan tidak kompetitif jarang berlaku; perencat tidak kompetitif yang biasa adalah bahan yang secara terbalik mengikat pada penambahan ES yang menimbulkan ESI:

Inhibisi dari kelebihan substrat kadang-kadang tidak kompetitif, kerana ini berlaku apabila molekul substrat kedua mengikat kompleks ES, sehingga menimbulkan kompleks ESS.
Sebaliknya, perencat tidak kompetitif hanya boleh mengikat pada enzim substrat seperti dalam kes sebelumnya: pengikatan substrat dengan enzim bebas mendorong pengubahsuaian konformasi yang menjadikan laman web ini dapat diakses oleh perencat.
Pemalar Michaelis Menten menurun apabila kepekatan perencat meningkat: nampaknya, pertalian enzim untuk substrat meningkat.
Protease serin
Mereka adalah sekumpulan enzim yang menjadi milik chymotrypsin dan trypsin.
Chymotrypsin adalah enzim proteolitik dan hidrolitik yang memotong di sebelah kanan asid amino hidrofobik dan aromatik.
Produk gen yang memberi kod untuk chymotrypsin tidak aktif (diaktifkan dengan arahan); bentuk chymotrypsin yang tidak aktif ditunjukkan oleh rantai polipeptida 245 asid amino. Chymotrypsin mempunyai bentuk globular kerana lima jambatan disulfida dan interaksi kecil lain (elektrostatik, daya Van der Waals, ikatan hidrogen, dll.).
Chymotrypsin dihasilkan oleh sel chymose pankreas di mana ia terkandung dalam membran khas dan dikeluarkan melalui saluran pankreas ke dalam usus, pada masa pencernaan makanan: chymotrypsin sebenarnya adalah enzim pencernaan. Protein dan nutrien yang kita makan melalui diet mengalami pencernaan untuk dikurangi menjadi rantai yang lebih kecil dan diserap dan diubah menjadi tenaga (mis. Amilase dan protease memecah nutrien menjadi glukosa dan asam amino yang sampai ke sel, melalui saluran darah mereka sampai ke vena portal dan dari sana dihantar ke hati di mana mereka menjalani rawatan lebih lanjut).
Enzim dihasilkan dalam bentuk tidak aktif dan diaktifkan hanya apabila mereka sampai di "laman web di mana mereka mesti beroperasi"; setelah tindakan mereka selesai, mereka akan dinyahaktifkan. Enzim, setelah dinyahaktifkan, tidak dapat diaktifkan semula: untuk mempunyai "tindakan pemangkin lebih lanjut, ia mesti diganti dengan" molekul enzim lain. Sekiranya chimitrypsin dihasilkan dalam bentuk aktif yang sudah ada di pankreas, ia akan menyerang yang terakhir: pankreatitis adalah patologi kerana enzim pencernaan yang sudah diaktifkan di pankreas (dan bukan di tempat yang diperlukan); beberapa di antaranya jika tidak dirawat tepat pada waktunya, membawa kepada kematian.
Dalam chymotrypsin dan dalam semua protease serin, tindakan pemangkin disebabkan oleh adanya anion alkohol (-CH2O-) di rantai sisi serin.
Protease serin mengambil nama ini tepat kerana tindakan pemangkinnya disebabkan oleh serine.
Setelah semua enzim melakukan tindakannya, sebelum dapat beroperasi semula pada substrat, ia mesti dipulihkan dengan air; "pelepasan" serin oleh air adalah tahap proses yang paling lambat, dan ini adalah fasa ini yang menentukan kelajuan pemangkinan.
Tindakan pemangkin berlaku dalam dua fasa:
- pembentukan anion dengan sifat pemangkin (anion alkoholat) dan serangan nukleofilik seterusnya pada karbon karbonil (C = O) dengan pembelahan ikatan peptida dan pembentukan ester;
- serangan air dengan pemulihan katalis (dapat melakukan tindakan pemangkinnya lagi).
Pelbagai enzim yang tergolong dalam keluarga protease serin dapat terdiri dari asam amino yang berlainan tetapi, untuk semuanya, laman pemangkin diwakili oleh anion alkohol dari rantai sisi serine.
Subfamily protease serin adalah enzim yang terlibat dalam pembekuan (yang terdiri dalam transformasi protein, dari bentuk tidak aktif mereka menjadi "bentuk lain yang aktif). Enzim ini memastikan bahawa pembekuan seefektif mungkin dan terhad dalam ruang dan masa (pembekuan mesti berlaku dengan cepat dan hanya berlaku di sekitar kawasan yang cedera). Enzim yang terlibat dalam pembekuan diaktifkan dalam lata (dari pengaktifan satu enzim, diperolehi berbilion enzim: setiap enzim diaktifkan , seterusnya mengaktifkan banyak enzim lain).
Trombosis adalah patologi disebabkan oleh kekurangan fungsi enzim pembekuan: ia disebabkan oleh pengaktifan, tanpa perlu (kerana tidak ada kecederaan), enzim yang digunakan dalam pembekuan.
Terdapat enzim modulasi (peraturan) dan enzim penghambat untuk enzim lain: berinteraksi dengan yang terakhir, mereka mengatur atau menghalang aktiviti mereka; malah produk enzim boleh menjadi penghambat enzim.Ada juga enzim yang berfungsi lebih banyak, semakin besar substrat yang ada.
Lysozyme
Luigi Pasteur mendapati, dengan bersin pada piring petri, bahawa di lendir terdapat enzim yang mampu membunuh bakteria: lisozim; dari bahasa Yunani: liso = ukuran apa; zimo = enzim.
Lysozyme mampu memecah dinding sel bakteria. Bakteria, dan organisma uniselular pada amnya, memerlukan struktur tahan mekanikal yang menghadkan bentuknya; di dalam bakteria terdapat tekanan osmotik yang sangat tinggi sehingga mereka menarik air. Membran plasma akan meletup jika tidak ada dinding sel yang menentang kemasukan air dan membatasi jumlah bakteria.
Dinding sel terdiri daripada rantai polisakarida di mana molekul N-asetil-glukosamin (NAG) dan molekul asid N-asetil-muramic (NAM) bergantian; ikatan antara NAG dan NAM dipecahkan oleh hidrolisis. Kumpulan karboksil NAM, di dinding sel, terlibat dalam ikatan peptida dengan asid amino.
Di antara pelbagai rantai, jambatan terbentuk yang terdiri daripada ikatan pseudo-peptida: percabangan disebabkan oleh molekul lisin; struktur secara keseluruhan sangat bercabang dan ini memberikan kestabilan yang tinggi.
Lysozyme adalah antibiotik (membunuh bakteria): ia berfungsi dengan membuat keretakan di dinding bakteria; apabila struktur ini (yang tahan mekanikal) pecah, bakteria menarik air sehingga pecah. Lysozyme berjaya memutuskan ikatan glukosid β-1,4 antara NAM dan NAG.
Tapak pemangkin lisozim diwakili oleh alur yang bergerak di sepanjang enzim di mana rantai polisakarida dimasukkan: enam cincin glukosidik rantai diletakkan di alur.
Di posisi tiga alur c "tersedak: di posisi ini hanya satu NAG yang dapat ditempatkan, kerana NAM, yang mempunyai dimensi yang lebih tinggi, tidak dapat masuk. Tapak pemangkin sebenarnya adalah antara kedudukan empat dan lima: kerana terdapat NAG di posisi tiga, pemotongan akan berlaku antara NAM dan NAG (dan bukan sebaliknya); oleh itu, pemotongan adalah spesifik.
PH optimum untuk kerja lisozim adalah lima. Di tempat pemangkin enzim, iaitu di antara kedudukan empat dan lima, terdapat rantai sisi asid aspartik dan asid glutamat.
Tahap homologi: mengukur kekeluargaan (iaitu persamaan) antara struktur protein.
Terdapat hubungan yang kuat antara lysozyme dan lactose-synthase.
Laktosa synthetase mensintesis laktosa (yang merupakan gula susu utama): laktosa adalah galaktosil glukosida di mana c "adalah ikatan glukosid β-1,4 antara galaktosa dan glukosa.
Oleh itu, laktosa synthetase mengkatalisis reaksi yang berlawanan dengan yang dikatalisis oleh lisozim (yang sebaliknya memisahkan ikatan glukosidik β-1,4)
Laktosa synthetase adalah dimer, iaitu, ia terdiri dari dua rantai protein, salah satunya mempunyai sifat pemangkin dan setanding dengan lisozim dan yang lain adalah subunit pengawalseliaan.
Semasa mengandung, glikoprotein disintesis oleh sel kelenjar susu dengan tindakan galatosil-tranferase (ia mempunyai "urutan homologi 40% dengan lisozim): enzim ini dapat memindahkan kumpulan galaktosil dari struktur tenaga tinggi ke struktur glikoprotein. Semasa kehamilan, ekspresi gen yang memberi kod galaktosisil-transferase disebabkan (terdapat juga ekspresi gen lain yang juga memberikan produk lain): terdapat peningkatan ukuran payudara kerana diaktifkan kelenjar susu (sebelum ini tidak aktif) yang mesti menghasilkan susu. Semasa melahirkan, α-laktalbumin dihasilkan yang merupakan protein pengawal: ia mampu mengatur kapasiti pemangkin galactosyl-transferase (dengan diskriminasi substrat). Galactosyl-transferase yang diubah oleh α-lactalalbumin mampu memindahkan galactosyl ke molekul glukosa: membentuk ikatan glikosid β-1,4 dan memberikan laktosa (laktosa synthetase).
Oleh itu, galaktosa transferase menyiapkan kelenjar susu sebelum melahirkan dan menghasilkan susu selepas bersalin.
Untuk menghasilkan glikoprotein, galactosyl transferase mengikat galactosyl dan NAG; semasa kelahiran lactal albumin mengikat galactosyltransferase menjadikan yang terakhir mengenali glukosa dan tidak lagi NAG memberikan laktosa.
.jpg)