Sekiranya sel berfungsi dalam keadaan anaerob, ia menghasilkan tenaga dengan menukar glukosa menjadi laktat dan, melalui kitaran Cori, membuang yang terakhir; jika ada oksigen yang tersedia (oleh itu dalam keadaan rehat), lebih daripada 90% glukosa dimakan secara aerobik dan hanya baki 10%, secara anaerob. Apabila terdapat keperluan untuk lebih banyak ATP daripada yang dapat disediakan oleh jalur aerobik (contohnya ketika otot berada dalam keadaan tertekan), maka bekalan tambahan diberikan oleh metabolisme anaerob (kita berada dalam keadaan kekurangan oksigen: sesak nafas, keletihan semasa bernafas, dan lain-lain): perlu untuk mempercepat metabolisme ini dengan menukar laktat (yang diperoleh dari glikolisis) menjadi glukosa melalui glukoneogenesis.
Metabolisme aerobik berkembang di mitokondria.
Enzim pertama yang ditemui dalam metabolisme aerobik adalah piruvat dehidrogenase; Lebih tepat untuk mengatakan bahawa piruvat dehydrogenase adalah kompleks enzim dan bukannya enzim, kerana ia adalah agregat 48-60 unit protein dengan tiga laman pemangkin bertindak berturut-turut.
Pyruvate dehydrogenase memangkinkan tindak balas berikut (redoks):
Pyruvate + NAD + + CoA-SH → Acetyl CoA + NADH + H + + CO2
CoA-SH adalah koenzim A: ia adalah turunan asid pantotenat; asetil koenzim A adalah thioester. Ini adalah proses redoks kerana karbon pertama piruvat beralih dari pengoksidaan nombor tiga hingga pengoksidaan nombor empat (ia teroksidasi) dan karbon kedua piruvat beralih dari pengoksidaan nombor dua ke pengoksidaan nombor tiga (ia teroksidasi). Kemudian piruvat dioksidakan (kehilangan dua elektron sama sekali) dan NAD dikurangkan.
Seperti disebutkan, piruvat dehydrogenase mempunyai tiga jenis aktiviti enzimatik, masing-masing disokong oleh kofaktor katalitiknya sendiri:
- tiamina pirofosfat (ia adalah turunan dari vitamin B1); ia aktif dalam bentuk deprotonated: karbanion terbentuk.
- lipoamide (ia adalah turunan asid lipoik); ia mengandungi jambatan disulfida yang sangat reaktif.
- flavin adenin dinucleotide (ia adalah turunan dari vitamin B2); ia adalah nukleotida dengan sifat redoks: pusat redoksnya terdiri daripada flavin.
Dalam sel eukariotik, metabolisme aerobik berlaku pada organel khusus sel yang merupakan mitokondria; pada bakteria metabolisme glukosa dan spesies lain berlaku di dalam sel tetapi tidak ada organel khusus.
Apabila piruvat memasuki mitokondria, ia dikenakan "tindakan karboksilase piruvat jika ada keperluan untuk melakukan glukoneogenesis (untuk menyusun semula bahan awal), atau ia boleh mengalami dehidrogenase piruvat jika perlu menghasilkan tenaga: "asetil koenzim A yang terbentuk oleh metabolisme aerobik merangsang tindakan piruvat karboksilase, oleh itu, ia mendorong gluokoneogenesis dan mengurangkan tindakan piruvat dehidrogenase.
Sekarang mari kita lihat bagaimana piruvat dehidrogenase berfungsi; pertama sekali, terdapat dekarboksilasi piruvat oleh tindakan pirofosfat tiamin.
Persekitaran yang berasid dapat menghambat metabolisme aerobik kerana bentuk anionik tiamina pirofosfat aktif yang akan diprotonasikan pada pH berasid dan dekarboksilasi tidak akan berlaku.
Dekarboksilasi adalah reaksi yang sukar kerana ikatan karbon-karbon mesti diputuskan; dalam kes ini, tindak balas termodinamik disukai oleh kenyataan bahawa perantaraan reaksi (hidroksietil-tiamin pyrofosfat) memberikan resonans (p-elektron molekul dilokalisasi): hidroksietil-tiamin pirofosfat wujud dalam tiga bentuk yang mungkin (resonans) dan ini menjadikannya agak stabil. Tambahan pula, hidroksietil-tiamin pirofosfat dalam bentuk anionik bertahan dalam jangka masa yang cukup lama untuk dapat berinteraksi dengan jambatan disulfida lipoamide (katalis pemangkin kedua piruvat dehidrogenase); jambatan disulfida adalah lengan berayun (ia terletak di hujung rantai fleksibel yang panjang) dan boleh berpindah dari satu tempat pemangkin ke tempat lain di kompleks enzim.
Kemudian lipoamida, melalui jambatan disulfida, mengikat hidroksietil-tiamin pirofosfat: asetil lipoamida diperolehi.Yang baru dijelaskan adalah langkah pertama tindak balas transasetilasi yang dikatalisis oleh enzim pertama kompleks piruvat dehidrogenase; pada fasa ini ikatan terputus antara kumpulan hidroksil dan tiamina pirofosfat yang kembali ke bentuk asalnya: tindak balas redoks berlaku di mana jambatan disulfida bertindak sebagai oksidan (dua atom sulfur dikurangkan) dalam perbandingan dalam kumpulan hidroksil yang dioksidakan menjadi asetil.
Selepas fasa ini, lengan osilasi lipoamide bergerak dan mendekati enzim kedua piruvat dehidrogenase yang menjalankan aktiviti transacetylase yang sebenarnya dengan membawa kumpulan asetil dengannya: fasa kedua reaksi transasetilasi yang dikatalisis oleh enzim kedua berlaku; dengan cara ini kita telah memperoleh asetil koenzim A. Kini perlu untuk memulihkan lipoamide yang dalam bentuk berkurang: enzim ketiga piruvat dehydrogenase campur tangan yang mengubah semula lipoamide dan memindahkan elektrodnya ke FAD yang dikurangkan menjadi FADH2. FAD / FADH2 dapat berfungsi sebagai pasangan redoks dalam dua tahap monoelektronik yang berbeza atau dalam satu tahap dua arah.
FADH2 segera memberikan elektronnya ke NAD + memperoleh FAD dan NADH + H +.
Asetil koenzim A, yang diperoleh seperti yang dijelaskan, adalah produk permulaan untuk kitaran Krebs (atau kitaran asid tricarboxylic).






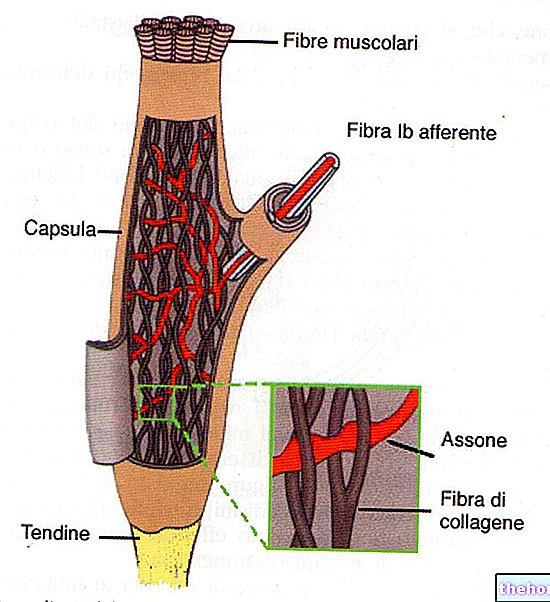


.jpg)