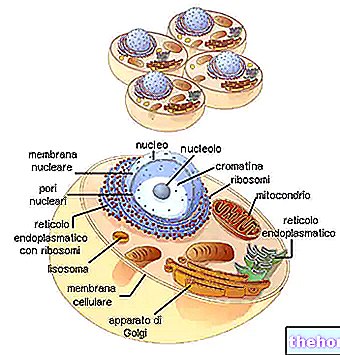
Untuk ada korespondensi antara maklumat polinukleotida dengan maklumat polipeptida, ada kod: kod genetik.
Ciri umum kod genetik boleh disenaraikan seperti berikut:
Kod genetik terdiri daripada kembar tiga, dan tanpa tanda baca dalaman (Crick & Brenner,).
Ia "diuraikan melalui penggunaan" sistem terjemahan sel terbuka "(Nirenberg & Matthaei, 1961; Nirenberg & Leder, 1964; Korana, 1964).
Ia sangat merosot (sinonim).
Penyusunan jadual kod tidak sengaja.
Kembar tiga "omong kosong".
Kod genetik adalah "standard", tetapi tidak "universal".
Melihat jadual kod genetik, harus diingat bahawa ia merujuk kepada terjemahan "RNAm ke polipeptida, yang mana asas nukleotida yang terlibat adalah A, U, G, C. Biosintesis rantai polipeptida adalah terjemahan dari urutan nukleotida dalam urutan asid amino.

Setiap triplet asas RNAm, yang disebut kodon, mempunyai pangkalan pertama di lajur kiri, yang kedua di baris atas, yang ketiga di lajur kanan. Mari kita contohi triptofan (iaitu Cuba) dan kita melihat bahawa kodon yang sesuai akan menjadi, dengan teratur, UGG. Sebenarnya, pangkalan pertama, U, merangkumi keseluruhan barisan kotak di bahagian atas; dalam ini, G mengenal pasti kotak paling kanan dan baris keempat kotak itu sendiri, di mana kita dapati cuba ditulis. Begitu juga, untuk mensintesis tetrapeptida Leucine-Alanine-Arginine-Serína (simbol Leu-Ala-Arg-Ser) kita dapat menemukan kodon UUA-AUC-AGA-UCA dalam kodnya.

Walau bagaimanapun, pada ketika ini, perlu diperhatikan bahawa semua asid amino tetrapeptida kita dikodkan (tidak seperti triptofan) oleh lebih daripada satu kodon. Bukan kebetulan bahawa dalam contoh yang baru saja dilaporkan, kami telah memilih kodon yang ditunjukkan. Kami mungkin mengekod tripeptida yang sama dengan urutan RNAm yang berbeza, seperti CUC-GCC-CGG-UCC.
Pada mulanya, fakta bahawa asid amino tunggal berhubungan dengan lebih dari tiga kembar diberi makna kebiasaan, juga dinyatakan dalam pilihan istilah degenerasi kod, yang digunakan untuk menentukan fenomena sinonim. Sebaliknya, beberapa data menunjukkan bahawa ketersediaan sinonim yang merujuk kepada kestabilan maklumat genetik yang berbeza sama sekali tidak sengaja. Ini nampaknya disahkan juga oleh penemuan nilai yang berbeza dari nisbah A + T / G + C dalam pelbagai tahap evolusi. Sebagai contoh, di prokariota, di mana keperluan untuk kebolehubahan tidak dipenuhi oleh peraturan Mendelisme dan neo-Mendelisme, nisbah A + T / G + C cenderung meningkat. Kestabilan yang lebih rendah akibatnya, dalam menghadapi mutasi, memberikan lebih besar peluang untuk kebolehubahan secara rawak dari mutasi gen.
Dalam eukariota, khususnya dalam sel multiselular, di mana sel organisma tunggal perlu menjaga warisan keturunan yang sama, nisbah A + T / G + C dalam DNA cenderung menurun, sehingga menurunkan peluang untuk mutasi gen somatik .
Kewujudan kodon sinonim dalam kod genetik menimbulkan masalah, yang telah disebutkan, mengenai banyaknya atau tidak antikodon dalam RNAt.
Sudah pasti bahawa terdapat sekurang-kurangnya satu RNAt untuk setiap asid amino, tetapi tidak pasti sama ada RNAt tunggal dapat mengikat satu kodon, atau dapat mengenali sinonim secara acuh tak acuh (terutama apabila ini hanya berbeza untuk asas ketiga).
Kita dapat menyimpulkan bahawa rata-rata terdapat tiga kodon sinonim untuk setiap asid amino, sementara antikodon sekurang-kurangnya satu, dan tidak lebih dari tiga.
Mengingat bahawa gen dimaksudkan sebagai rangkaian tunggal urutan DNA polinukleotida yang sangat panjang, jelas bahawa awal dan akhir gen tunggal semestinya terkandung dalam ingatan.
BIOSYNTHESIS PROTEIN
Di bahagian DNA yang berlainan terdapat pembukaan rantai berganda dan sintesis pelbagai jenis RNA.
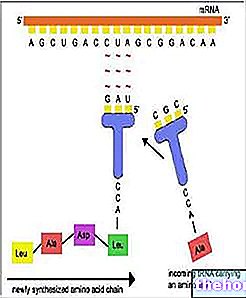
Semasa langkah pemuatan, RNAt mengikat asid amino (sebelumnya diaktifkan oleh ATP dan oleh enzim tertentu). "Mesin" biosintetik tidak dapat "membetulkan" tRNA yang dimuat secara tidak betul.
RNAr kemudian berpecah menjadi dua subunit dan, dengan mengikat protein ribosom, menimbulkan pemasangan ribosom.
RNAm, melewati sitoplasma, mengikat ke ribosom, membentuk polysome.Setiap ribosom, yang mengalir pada utusan, secara beransur-ansur menjadi tuan rumah RNAt yang melengkapi kodon relatif, mengambil asid amino dan mengikatnya ke rantai polipeptida dalam pembentukan.
RNAt yang stabil kembali memasuki peredaran. Ribosom juga digunakan lagi, melepaskan polipeptida yang sudah dipasang.
Utusan, kurang stabil kerana semuanya monocatenary, dibelah (oleh ribonuclease) ke dalam ribonukleotida konstituen.
Oleh itu kitaran berterusan, mensintesis satu demi satu polipeptida pada RNA pemesejan yang dibekalkan oleh transkripsi.